Mouches volantes - Glaskörpertrübung oder Nervensystem?
Archivmeldung vom 22.12.2009
Bitte beachten Sie, dass die Meldung den Stand der Dinge zum Zeitpunkt ihrer Veröffentlichung am 22.12.2009 wiedergibt. Eventuelle in der Zwischenzeit veränderte Sachverhalte bleiben daher unberücksichtigt.
Freigeschaltet durch Thorsten Schmitt![Vereinfachte schematische Darstellung der Beziehung der verschiedenen Nervenzellen in den Netzhautschichten. Quelle: Wörterbuch Biologie, hg. v. Gertrud Scherf, München: Deutscher Taschenbuch Verlag 1997 [Elektronische Ausgabe: Digitale Bibliothek, Bd. 140, Berlin: Directmedia 2006]](https://www.extremnews.com/images/article_landscape-213df31d95c599d79578994e398dd552.png)
Seit Jahrhunderten versuchen Gelehrte, eine Erklärung für die beweglichen Punkte und Fäden im Gesichtsfeld zu finden. Schon früh dachte man sich die Ursache im Auge und verstand das Phänomen als eine Störung oder Degeneration irgendwo zwischen Pupille und Netzhaut – heute gelten Mouches volantes als Trübung des Glaskörpers. Genaue Beobachtungen aber enthüllen geordnete Strukturen und lassen die Degenerationsthese fragwürdig erscheinen.
Im Nervensystem des Sehens befinden sich ähnliche Strukturen – was zur Vermutung führt, dass die fliegenden Mücken dort ihren Ursprung haben.
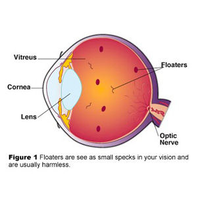
In der Augenheilkunde ist „Mouches volantes“ (MV) ein Sammelbegriff für Glaskörpertrübungen, die auf unterschiedliche Ursachen zurückgeführt werden. Meistens jedoch gilt die Erscheinung als normale altersbedingte Trübung des Glaskörpers infolge der Verflüssigung (Synchisis) und der oft damit verbundenen Kollabierung der Kollagen-Hyaluron-Struktur des Glaskörpers (Syneresis), sowie seiner Abhebung von der Netzhaut. Diese von der Medizin als harmlos bezeichneten MV sind bei Tageslicht im Blickfeld als bewegliche, transparente vereinzelte Punkte und Fäden wahrnehmbar.
Diese medizinische Wahrnehmung der MV als „Störung“ oder heute eben „Glaskörpertrübung“ beruht meiner Ansicht nach auf einer oberflächlichen Beobachtung des Phänomens und trifft nicht zu. Ich vertrete den Standpunkt, dass MV Ausdruck eines polaren Ordnungsprinzips sind, das sowohl kulturelle wie spirituelle Bedeutung hat, das sich aber auch in den Eigenschaften des visuellen Nervensystems finden lässt. Um diese Behauptung zu erläutern gehe ich vom beobachteten Phänomen aus und suche nach strukturellen Ähnlichkeiten in der Physiologie des Sehens. Dabei nehme ich mit den Gestaltpsychologen an, dass die in der Wahrnehmungserfahrung (und m.E. auch in geistigen bzw. Bewusstseinsregungen) gegebene phänomenale Ordnung das Ergebnis von dynamischen neurologischen Ordnungsprozessen im Kortex ist (psychophysisches Isomorphieprinzip). Mouches volantes wären demnach ein Bindeglied zwischen Physiologie, Neurologie, Psychologie, Parapsychologie, Philosophie, Religion und Spiritualität.
Mouches volantes als dualistisch-polare Kugeln und Fäden
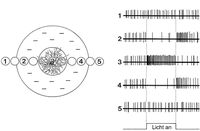
Ich beginne mit simplen Beobachtungen: Mouches volantes sind Punkte und Fäden. Die Fäden sind dabei mit aneinander gereihten Punkten gefüllt, die deutlicher oder weniger deutlich sichtbar sind. Die Punkte sind kreisrund und konzentrisch, verfügen über einen Kern und ein Umfeld, d.h. sie sind polar. Zur Polarität gesellt sich ein Dualismus, denn es gibt zwei Arten von Punkten: Solche mit hellem Umkreis und dunklem Kern, und solche mit dunklem Umkreis und hellem Kern. Wir können also von einem dualistisch-polaren Prinzip in den MV sprechen. Es ist schwer vorstellbar, dass zufällig verklumpte Glaskörperfibrillen so genaue und repetierbare morphologische Eigenschaften aufweisen.
Mouches volantes als rezeptive Felder
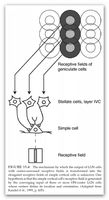
Gibt es im visuellen Nervensystem nun Entsprechungen dieser Eigenschaften der MV? Tatsächlich können wir in der Netzhaut und im Verarbeitungsprozess der neuronalen Signale in den Sehbahnen dasselbe dualistisch-polare Prinzip finden: nämlich in der Organisation der Nervenzellen der Netzhaut, der Sehbahnen und der visuellen Hirnarealen (Cortices).
Wenn Licht in das Auge fällt, durchwandert es verschiedene Zellschichten der Netzhaut und wird in der hintersten Schicht von der ersten Art von Neuronen oder Nervenzellen aufgenommen, nämlich von den Photorezeptoren (Stäbchen und Zapfen). Sie haben die Aufgabe, Lichtimpulse in elektrische Signale umzuwandeln und an die zweite Art von Nervenzellen, die Bipolarzellen, weiterzugeben. Diese geben ihre Information wiederum an den dritten Neuronentyp weiter, die Ganglienzellen. Im Durchschnitt nimmt jede Ganglienzelle die Informationen von ca. 100 Stäbchen oder von 4-6 Zapfen entgegen. Photorezeptoren, Bipolar- und Ganglienzellen werden durch zwei weitere Neuronen miteinander verbunden, nämlich durch Amakrinzellen und Horizontalzellen, die für den lateralen (seitlichen) Informationsaustausch unter den Neuronen sorgen und auf diese Weise den Wahrnehmungsvorgang koordinieren, modulieren und integrieren.
Der Weg der Lichtenergie ist aber damit nicht zu Ende: Die Ganglienzellen der Netzhaut feuern bei genügend „Zündstoff“ (Aktionspotential) durch ihre weiterleitenden Fortsätze (Axone) durch den Sehnerv bis in den Äußeren Kniehöcker (Corpus geniculatum laterale) im Hirn. Im Kniehöcker werden die visuellen Inputs an die vierte Art von Neuronen, die verschiedenen kortikalen Nervenzellen, übertragen, die sich in den visuellen Zentren des Gehirns finden lassen. Die Informationen werden über Nervenfaserbündel v.a. an den so genannten primären visuellen Kortex (V1) gesendet, dann auch an andere visuelle Zentren.
Für eine neurologische Interpretation der MV ist nun interessant zu wissen, dass jede Nervenzelle nach den Photorezeptoren ein so genanntes „rezeptives Feld“ hat. Damit ist ein definierter Bereich im Sehfeld gemeint, aus dem Nervenzellen Lichtsignale erhalten. Jedes Neuron antwortet also auf Stimuli, die von Stäbchen oder Zapfen stammen, die innerhalb dieses rezeptiven Feldes liegen. Das Besondere an diesen Feldern ist ihre Form und ihre polare Organisation: Rezeptive Felder von Bipolar- und Ganglienzellen sind kreisrunde konzentrische Felder, die sich durch ein Zentrum und eine Umgebung auszeichnen. In Fachkreisen spricht man von einem „center-surround antagonistic receptive field“ (CSARF). Das Zentrum entspricht dabei ungefähr dem dendritischen Radius der Nervenzelle.
Dabei gibt es zwei Arten von Neuronen, die sich funktional und auch morphologisch unterscheiden lassen: solche, die auf die Beleuchtung des Zentrums ihres rezeptiven Feldes reagieren (On-Zentrum) und solche, die auf die Beleuchtung ihres Umfeldes reagieren (On-Peripherie bzw. Off-Zentrum). Werden Photorezeptoren im Zentrum eines rezeptiven Feldes einer On-Zentrum-Bipolarzelle beleuchtet, lösen sie eine Erregung (Depolarisation) an der Zellmembran aus; die Zelle gibt diese Information an die entsprechende On-Ganglienzelle weiter, bei der sich wiederum die Entladungsrate (Aktionspotentiale pro Zeiteinheit) erhöht. Wird dagegen das Umfeld der On-Bipolarzelle beleuchtet, wird die Zelle gehemmt (Hyperpolarisation), d.h. die Entladungsrate verringert sich. In der Off-Zentrum-Bipolarzelle verhält es sich genau umgekehrt: Die Beleuchtung des Zentrums bewirkt eine Hyperpolarisation der Zellmembran und hemmt die Zelle, während sie durch die Beleuchtung des Umfelds depolarisiert, d.h. erregt wird; in der entsprechenden Ganglienzelle wird die Entladungsrate im ersten Fall verringert, im zweiten erhöht. Sowohl Bipolar- wie Ganglienzellen enthalten ihre Inputs nicht nur von den Photorezeptoren (vertikaler Weg), sondern auch lateral von Amakrin- und Horizontalzellen (horizontaler Weg). Diese vermitteln Signale im Umfeld eines rezeptiven Feldes, d.h. außerhalb des dendritischen Baumes einer Nervenzelle, und bewirken auf diese Weise die jeweils gegenteilige Reaktion der Nervenzelle bei der peripheren Beleuchtung. Sie sind somit Bestandteil des Zentrum-Umgebungs-Antagonismus. Diese Art der Signalverarbeitung, die einen klar definierten Bereich erregt, die umgebenden Zellen aber hemmt, trägt zur Kontrastierung unseres Sehens bei: Auf diese Weise wird scharfes Sehen und die klare Unterscheidung zwischen dunklen und hellen Feldern möglich. Was Ganglienzellen also an die kortikalen Sehzentren weiterleiten, sind nicht die Stimulationen einzelner Photorezeptoren, sondern die Codes von rezeptiven Feldern, die räumlich summierten Erregungs- und Hemmungsprozesse, die in diesen Feldern ausgelöst wurden.
Die rezeptiven Felder der Sehnervenzellen weisen somit genau dasselbe dualistisch-polare Prinzip auf wie die Kugeln der Mouches volantes. Was aber ist mit den Fäden? Auch hier gibt es eine Entsprechung, denn in bestimmten kortikalen Bereichen sind die rezeptiven Felder der Neuronen nicht kreisrund und konzentrisch, sondern länglich. Es sind Streifen mit verschiedenartiger räumlicher Ausrichtung, d.h. diese Neuronen reagieren sehr deutlich auf Lichtstimuli, die sich durch lineare Eigenschaften auszeichnen (Linien, Spalte, Streifen etc.); zudem sind sie auf bestimmte Winkel und Ausrichtungen dieser Linien getrimmt („orientation selecitivty“). Allerdings versteht man noch nicht genau, wie der Output der Neuronen im Kniehöcker, welche ausschließlich nach dem Prinzip der Zentrum-Umgebung organisiert sind, in die „orientation selectivity“ der V1-Neuronen übersetzt wird. Eine heute breit akzeptierte und einleuchtende Hypothese ist, dass die länglichen rezeptiven Felder im Kortex durch die verschmelzenden bzw. überlappenden Inputs von Zellen des Äußeren Kniehöckers zustande kommen; jene Zellen haben kreisrunde rezeptive Felder und enden im selben rezeptiven Feld einer entsprechenden V1-Nervenzelle.
Auch die Fäden der MV, die aus aneinander gereihten Kugeln bestehen, könnten also als rezeptive Felder von Neuronen in kortikalen visuellen Sehzentren aufgefasst werden. Dies würde insofern Sinn machen, als die Fäden bereits eine komplexere Struktur aufweisen als die Kugeln, und die Repräsentation von komplexeren Formen allgemein den Neuronen in höheren visuellen Zentren zugeschrieben wird.
Mögliche Einwände: Die Wahrnehmung rezeptiver Felder, die Beweglichkeit der Mouches volantes und die Vitrektomie
Mouches volantes wären demnach nicht die chaotische „Störung“ im Glaskörper, als die sie heute angenommen werden, sondern ein natürlicher Teil des neurophysiologischen Systems. Eine solche Behauptung steht im Widerspruch zur heute wissenschaftlich akzeptierten Vorstellung über MV. Insbesondere folgende Fragen wären zu klären: Wenn MV der visuelle Ausdruck von rezeptiven Feldern sind – warum können wir überhaupt rezeptive Felder sehen? Und warum können wir nur vereinzelte davon sehen, und nicht dieselbe Fülle, die wir in der Netzhaut antreffen? Eine weitere Frage betrifft die Bewegung von MV: Warum erfahren wir die MV in unserem Gesichtsfeld als so beweglich, z.T. durch unseren Blick beeinflussbar, z.T. eine Eigenbewegung aufweisend – wenn es sich dabei um unbewegliche rezeptive Felder in der Netzhaut handelt? Und schliesslich: Wie kann die Ursache der MV im neuronalen visuellen System liegen, wenn Augenärzte ihre Patienten von MV durch die Vitrektomie (Glaskörperentfernung) befreien?
Allen diesen Einwänden kann – zumindest auf theoretischer Ebene – begegnet werden. Einige zentrale Punkte sollen hier genannt werden: Es ist in der Wissenschaft noch immer ein Rätsel, wie und wo genau das Licht, das von der Retina empfangen wird, in visuelles Bewusstsein übersetzt wird. Klar ist, dass visuelle Erfahrungen Konstruktionen des Nervensystems sind, die sich nicht nur aus äußeren Stimuli speisen, sondern auch durch innere Reize zustande kommen. Von daher ist es denkbar, dass der visuelle Output davon abhängt, auf welcher Stufe der neuronalen Verarbeitung die bewusste Wahrnehmung einsetzt. Unsere vertraute materielle Welt ist demnach das Resultat einer Bewusstwerdung in den höheren visuellen Zentren, wo die neuronalen Reize vielfach miteinander verrechnet worden sind. Die Wahrnehmung von Mouches volantes dagegen könnte entsprechend durch eine Bewusstwerdung der neuronalen Reize auf hierarchisch tieferen bzw. früheren Verarbeitungsstufen zustande kommen, etwa im Äußeren Kniehöcker, im Sehnerv und sogar in der Netzhaut. Die Erregung von rezeptiven Feldern ist hier noch nicht zu komplexen Bildern verarbeitet, sondern liefert relativ isolierte, nicht in den individuellen Erfahrungshintergrund integrierte Eigenschaften wie Form, Farbe, Bewegung sowie Licht-und-Schatten-Kontraste.
Neben den äußeren Lichtstimuli bräuchte es weiterhin die Annahme eines Bewusstseinsprinzips, das die Signale der Nervenzellen reguliert, modifiziert und integriert. Ein solches Bewusstseinsprinzip, das seine materielle Grundlage im dynamischen Wechselspiel von neuronalen Feedforwards und Feedbacks hat, entscheidet, ob ein Nervenimpuls als rezeptives Feld bzw. als MV ins Bewusstsein gelangt, oder ob erst das komplexe verrechnete Resultat vieler Nervenimpulse wahrgenommen wird. Bewusstsein vermittelt also seine Struktur in der spezifischen Aktivierung rezeptiver Felder von Nervenzellen auf frühen Verarbeitungsstufen; diese Struktur zeigt sich uns visuell in unserem individuellen Muster der MV. Hier erhält die Beobachtung der eigenen MV insofern eine spirituelle Bedeutung, als es sich dabei um das Ergründen des eigenen Bewusstseins handelt.
Das Bewusstseinsprinzip liefert auch einen Ansatz, um die Bewegungen der MV zu erklären, zusammen mit dem psychologischen Konzept der „Scheinbewegung“ (apparent motion/movement). Dieses bezeichnet Bewegungseindrücke, die nicht durch bewegte Umweltreize zustande kommen, sondern auf den Anblick von stationären Reizen zurückgehen, die in wechselnden Folgen gezeigt werden. Beispiel Kino: Das Bild, das auf die Kinoleinwand projiziert wird, macht einen bewegten Eindruck – obwohl wir wissen, dass nur unbewegte Einzelbilder projiziert werden, und dass unser Hirn diese schnelle Abfolge von Einzelbildern zu einem bewegten Bild verrechnet. So ist es vorstellbar, dass MV durch die Verrechnung von sukzessiver Reizung unterschiedlicher Netzhaut- und kortikaler Regionen als beweglich erscheinen. In unserer Theorie kann diese Reizung allerdings nicht von äusseren Stimuli herrühren, sondern vom Bewusstseinsprinzip, das neurophysiologisch ebenfalls anregende oder hemmende Signale im Nervensystem bewirkt. Auch die scheinbare Abhängigkeit der MV-Bewegungen durch Augenbewegungen widerspricht dem nicht, insofern körperliche Bewegungen m.E. vom verursachenden Bewusstseinsprinzip nicht zu trennen sind: Unsere körperliche Aktivität ist ein Resultat unseres Bewusstseinszustandes; umgekehrt erregen körperliche und geistige Bewegungen wie Augenbewegungen, Konzentration, Emotionen etc. unser Nervensystem, wirken bewusstseinstransformativ und können auf diese Weise die Wahrnehmung von Bewegungen bzw. Scheinbewegungen hervorbringen. Das bedeutet, dass wir nicht einfach nur die MV mit unseren Augen bewegen, sondern dass sich unsere Augen auch gemäss dem visuell gewordenen dynamischen Ausdruck unseres Bewusstseins bewegen. Es bedeutet, dass wir selbst das Fliessen der MV verursachen oder auch stoppen können – durch unsere körperlichen und geistigen Bewegungen und unsere Erfahrung, d.h. durch unser Bewusstsein, welches die entsprechenden Neuronen aktiviert und deren rezeptiven Felder wir als MV sehen.
Das auf den ersten Blick schlagkräftigste Argument gegen die Vorstellung, dass MV auf eine spezifische Aktivierung von Nervenzellen mit entsprechenden rezeptiven Feldeigenschaften zurückzuführen sind, ist die Vitrektomie, heute für Glaskörpertrübungen meistens in der Variante der effektiveren Floaterektomie durchgeführt. Durch diese chirurgische Entfernung von Glaskörperpartien sollen die angeblich im Glaskörper befindlichen MV ebenfalls beseitigt werden. Es gibt dazu klinische Studien und Statistiken, die den Erfolg der Floaterektomie bescheinigen, obwohl die „störenden Trübungen“ nicht in jedem Fall beseitig werden können, wie Patientenberichten zuweilen zu entnehmen ist.
Zentral scheint mir jedoch, dass in erfolgreichen Fällen selten klar ist, um welche Arten von MV es sich dabei gehandelt hat. Tatsächlich vorhandene Einlagerungen irgendwelcher Art können zwar objektiv festgestellt und entfernt werden. Häufig jedoch kann der Augenarzt keine Trübungen des Auges beim Patienten feststellen. Insistiert der Patient, wird oft trotzdem eine Floaterektomie durchgeführt, wobei dann eher der subjektiven Leidensdruck des Patienten als die MV behandelt werden. Möglicherweise handelt es sich bei erfolgreichen Floaterektomien also nicht um diejenigen MV, über die ich hier schreibe.
Wo es sich bei einer erfolgreichen Floaterektomie um dieselben MV handelt (was in jedem Einzelfall zu prüfen wäre), wäre ein anderer Gedanke in Betracht zu ziehen: Möglicherweise verändert eine Floaterektomie durch die Entfernung des kollagen- und hyaluronhaltigen Glaskörpergerüstes die Lichtstrahlungsverhältnisse im Glaskörper. Das Licht würde also nicht mehr in derselben Weise (z.B. in derselben örtlichen Dichte) auf die Netzhaut geleitet. Und das heißt, dass die Nervenzellen der Netzhaut nicht mehr in derselben Weise gereizt werden wie vor der Operation. Diese veränderte äussere Reizung würde zudem auch eine veränderte Aktivität des Bewusstseinsprinzips mit sich bringen. Dies könnte erklären, weshalb MV, als eine visuelle Erscheinung der neuronalen Tätigkeit im Sehsystem, nach einer Floaterektomie vom Patienten nicht mehr oder verändert gesehen werden. Dies wäre genauer abzuklären. Theoretisch jedenfalls ist die Floaterektomie kein zwingendes Gegenargument für die Annahme, dass MV durch die Funktion rezeptiver Felder entstehen.
Diese Arbeit kann die gemachten Behauptungen nicht belegen, viele Fragen bleiben offen. Der Text soll in erster Linie neue Impulse für die weitergehende Erforschung der MV geben und zu unkonventionellen Interpretationen unter Berücksichtigung der Beobachtenden ermutigen. Gerade der Einbezug des Subjekts eröffnet den Raum für verschiedene Interpretationen und Erklärungen, nicht nur ophthalmologische, sondern auch psychologische, historische, anthropologische und spirituelle – ein Schritt in die Richtung einer ganzheitlichen Augenheilkunde, die die klassischen Ophthalmologie sinnvoll ergänzen könnte.
Quelle: Floco Tausin
Anmerkung der ExtremNews Redaktion:
Floco Tausin ist Autor des Buches "Mouches volantes. Die Leuchtstruktur des Bewusstseins" (Buchbesprechung hier). Der Name Floco Tausin ist ein Pseudonym. Der Autor studierte an der geisteswissenschaftlichen Fakultät der Universität Bern und befasst sich in Theorie und Praxis mit der Erforschung subjektiver visueller Phänomene im Zusammenhang mit veränderten Bewusstseinszuständen und Bewusstseinsentwicklung. 2004 veröffentlichte er die mystische Geschichte „Mouches Volantes“ über die Lehre des im Schweizer Emmental lebenden Sehers Nestor und die spirituelle Bedeutung der Mouches volantes. In diesem Buch berichtet er über die den alten Griechen bereits bekannten, von heutigen Augenärzten als harmlose Glaskörpertrübung betrachtet und für viele Betroffene ärgerlich: Mouches volantes, Punkte und Fäden, die in unserem Blickfeld schwimmen und bei hellen Lichtverhältnissen sichtbar werden.
Die Erkenntnis eines im schweizerischen Emmental lebenden Sehers stellt die heutige Ansicht radikal in Frage: Mouches volantes sind erste Teile einer durch unser Bewusstsein gebildeten Leuchtstruktur. Das Eingehen in diese erlaubt dem Seher, über den Tod hinaus bewusst zu bleiben.
Mouches volantes: Glaskörpertrübung oder Bewusstseinsstruktur? Eine mystische Geschichte über die nahe (f)liegendste Sache der Welt.